Deep Learning: The Cerebellar Circuit
Introduction
Life is sure convenient, isn’t it? There are an innumerable number of different actions we perform every day without even giving it a second thought. Walking, talking, writing and so on are all examples of these actions, ones that seem mundane at the surface but are truly incredibly complex. For example, walking, requires the activation of numerous cortices and nuclei of the brain, such as the prefrontal cortex for containing the cognitive processes which initiate the action, the premotor cortex which acts as the bridge between these processes and the action they cause, and finally the motor cortex which converts all this neural activity to the format of a complex cognitive map of all your body’s muscles and which specific ones need to contract in order to perform the action (Lim). It is the final step, however, that is the most complex and undeniably important, coordination. This vast majority of this step takes place in the relatively small and seldom known region of the brain, the pons, the middle third of the brainstem, or more specifically, its bizarre oblong outgrowth, the cerebellum.
Quick History: Evolution and Discovery
The cerebellum is evolutionarily among the oldest regions of the vertebrate brain. It evolved nearly half a billion years ago in the vaguely fish-like ancestor of vertebrates from the nuclei on the dorsal side of the pons, which are closely associated with regulating bodily functions and facial reflexes. All it took was a simple invagination of these nuclei into the surrounding tissue and the cerebellum came into being (Hodos). 550 million years and many evolutionary cerebellar enhancements later, we arrive at its initial discovery in the late 18th century by Luigi Ronaldo. He was the first person to connect motor impairment with lesions to the cerebellum, giving humanity its first key insight into its true function. It was not until nearly a century later that Jan Evangelista Purkinje, through experimental microscopy methods, first described that the brain and nervous system was composed of individual microscopic units known as neurons in 1839. The neurons he specifically discovered with purkinje cells, the giant cells which compose the bulk mass of the cerebellar cortex and will be thoroughly discussed in the following section. At last, we arrive at our final destination, for the purposes of this exploration at least, where Janos Szentagothai and John Eccles through morphological and electrophysical analysis, were able to outline the internal anatomy and circuitry of the cerebellum in the 1960s (Glickstein), which will be the subject of the explanatory section of this report.
The Science: Where Biology Ends and Physics Begins
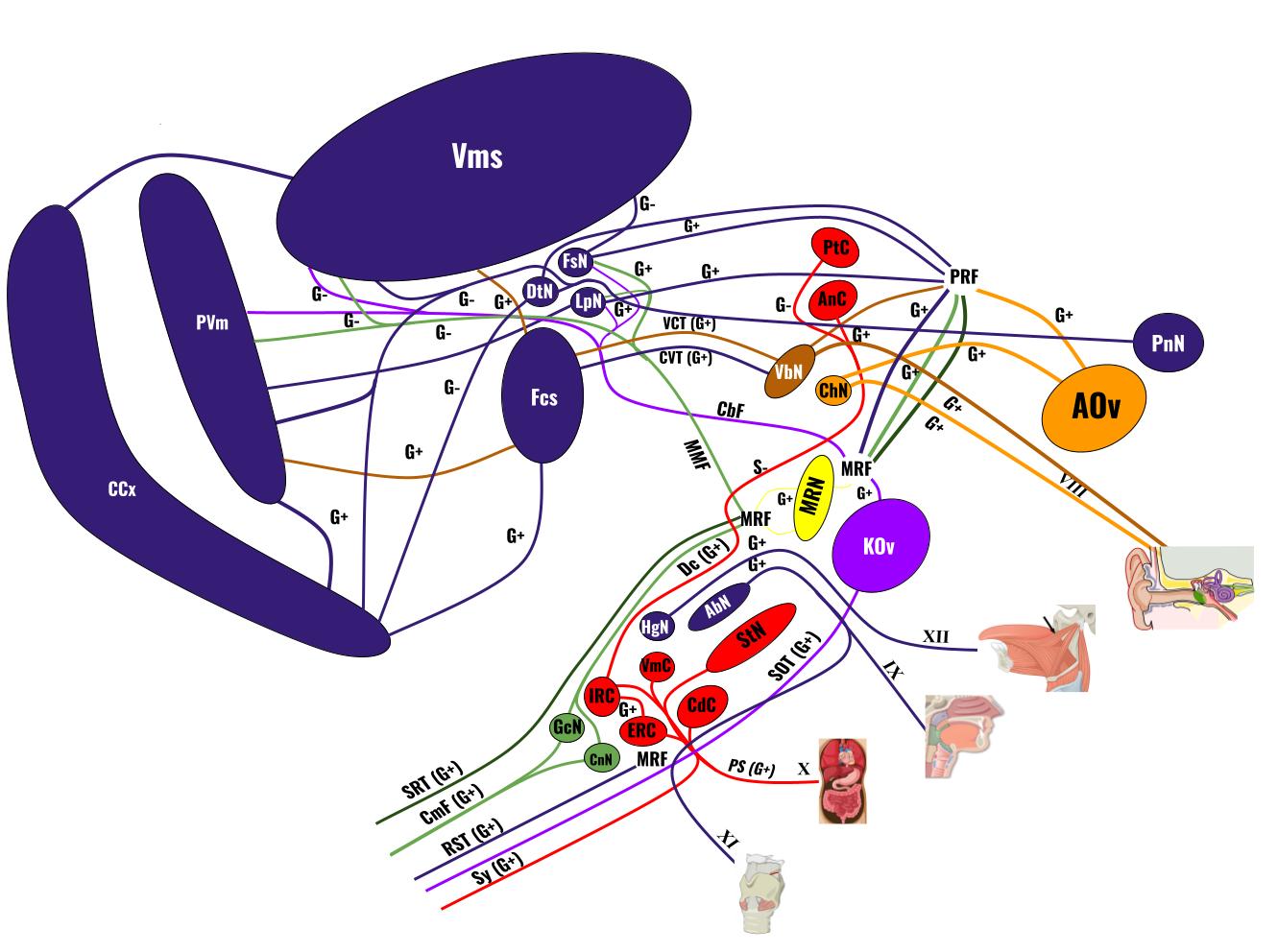
The cerebellum consists of four main regions, the ancient vestibulocerebellum which fixes gaze, head position and posture, the slightly newer deep cerebellar nuclei which transmit signals between the other two cerebellar structures and the rest of the brain, the fairly new spinocerebellum which helps with basic motor coordination, and the new, prized cortex of the cerebellum, which stores the procedural memory for every single action in its sprawling, fleshy, folded labyrinth. Compared to its far better known counterpart, the cerebral cortex, the cerebellar cortex is far simpler in structure, possessing only three layers, the granular, purkinje and molecular layers, each dedicated to a different cell type. The granular cells which compose the bulk of the cells in the cerebellum, all reside in the granular layer and project their axons into the molecular layer in what are called parallel fibers, because they all run parallel to each other, never crossing or touching. Perpendicular to these fibers are the tree-like dendrites of the purkinje cells which synapse with them in the molecular layer, whilst having their bodies in the purkinje layer, and projecting down past the granular layer toward the deep cerebellar nuclei. Granular cells receive information from neural fibers known as mossy fibers, which consist of kinesthetic information from the spinal cord and motor signals from the motor cortex (remember this from earlier?). On the other hand, purkinje cells receive inputs from both granular cells’ parallel fibers, as mentioned, and climbing fibers, which originate from the inferior olivary nucleus, an important kinesthetic-processing center in the brain. All of these excitatory signals then lead to a single inhibitory response from a purkinje cell to the deep cerebellar nuclei. See the visual aid below for a better understanding (ignore the inhibitory cells, such as the basket and stellate cells as they only play a minor role in cerebellar processes):
Kano, Masanobu, “Simplified Scheme of Cerebellar Neural Circuitry.” Research Gate.
How does this all relate to physics? This entire neural circuit essentially equates to a NOT logic gate. Whatever excitatory signal enters the circuit from the mossy and climbing fibers is converted into an inhibitory by the purkinje cell to inhibit the activity of the deep cerebellar nuclei. To complete this circuit, the deep cerebellar nuclei (DCN) output back to the motor cortex, from where many of the mossy fibers originate in the first place. Every time a movement occurs, it stimulates a mossy fiber signal which is then inverted by this NOT gate into a signal which prevents the DCN from sending a signal back to the motor cortex to stimulate its activity.
Now, to fully understand this circuit, we must delve into how it can be altered. Whenever granule cells/mossy fibers stimulate a purkinje cell, it creates a single inhibitory response, known as a simple spike. However, when a climbing fiber stimulates a purkinje cell, due to how interconnected it is with its dendrites, it creates a powerful, long-lasting inhibitory response, known as a complex spike. After the spike, it enters a refractory period in which it cannot fire. This contrasts simple from complex spikes. Repeated stimulation by both parallel/mossy fibers and climbing fibers will cause a purkinje cell to neurochemically alter its dendritic structure to disconnect parallel fibers and solely receive inputs from climbing fibers, a process known as long term depression. A depressed purkinje cell, unlike its prior state, can only inhibit DCN activity through complex spikes. According to the Marr-Albus-Ito model of cerebellar function, the most popular and widely accepted one, purkinje cells and by extension, the cerebellum, receive body awareness, position and balance information primarily from the mossy/parallel fibers, causing them to constantly fire in simple spikes to inhibit DCN/motor activity. The DCN functions according to a similar but opposite basis, sending a signal to the motor cortex for every proprioceptive or motor signal it receives from the mossy fibers. Purkinje cells can essentially be thought of as filters, selectively inhibiting DCN activity depending on whether they are depressed or not. If an error in movement occurs, such as a missed free throw in basketball, there will be a sudden burst of activity in the involuntary kinesthetic pathways leading from the spinal cord to the inferior olivary nucleus and into its climbing fibers, causing the corresponding purkinje cells (the ones linked to the motor neurons that caused the error as well as the proprioceptive pathway that sensed it) to output a complex burst to the DCN, inhibiting their firing in the short period that follows, thereby correcting the error temporarily. If this error repeats enough, then through long term depression, the purkinje cells causing it will lose their connection to the parallel fibers and no longer will be able to respond to kinesthetic or motor information through short spikes. The structure of the cerebellum will thus be permanently altered so as to never repeat those errors again. This is the crux of the phrase, “practice makes perfect”. Every time an action is repeated, this whole process repeats along with it, altering the cerebellum’s structure in such a way as to repeatedly refine and better the action by eliminating the firing capabilities of error-prone purkinje cells and their pathways (Purves). The final product is the reward reaped from dedicating the time to rewire one’s cerebellum, a new skill acquired.
Current Trends: Deep Learning
It was this complex yet simple self-altering circuit (as well as many others like it in the brain) that inspired machine learning in the first place. More specifically, one of the most prominent forms of machine learning, known as deep learning, was developed to replicate this process. Just as the cerebellum alters its inhibitive behaviors in response to depressive error signals, AI’s being trained through deep learning are exposed to the specific features of a dataset they are intended to identify. Both systems through repeated exposure to information, adapt to process it as intended by tweaking their parameters and internal configuration. The one substantial difference is that the cerebellum can rewrite its own parameters while the AI may need guidance from developers, but other than that, it is completely autonomous (What Is Machine Learning? | How It Works, Techniques & Applications).
The Future: Artificial Intelligence
How exactly is deep learning applicable to our modern world? It is completely transforming learning and our classrooms at this very moment. There is a relatively little known AI system, one that barely any of us have probably heard of, known as ChatGPT which uses deep learning as its foundation. The more data it is fed by developers, users and the worldwide web, the better it is able to adapt to the task it was given to fulfill. This is the main downside of deep learning, the immense amount of data that is required for it to function effectively. That is also, not coincidentally, the same downside to cerebellar motor learning, the immense amount of data that the system must be fed through repetition of desired movements or in other words practice. Moving on into the future, deep learning will continue to be employed with greater efficiency, requiring less data input and time, to contribute to the forefront of artificial intelligence (Singla).
Conclusion
To say the cerebellum is an amazing brain formation and adaptive learning system is an understatement. Not only is it responsible for the fine-tuning of each and every action we take, but it is in small thanks to attempts to imitate its unparalleled efficiency, that our lives and education are being forever changed by this unprecedented AI revolution we live in the midst of.
Works Cited
Lim, Shannon B., et al. “Brain Activity during Real-Time Walking and with Walking Interventions after Stroke: A Systematic Review.” Journal of NeuroEngineering and Rehabilitation, vol. 18, no. 1, 15 Jan. 2021, https://doi.org/10.1186/s12984-020-00797-w. Accessed 22 Apr. 2021.
Hodos, William. “Evolution of Cerebellum.” Encyclopedia of Neuroscience, pp. 1240–1243, https://doi.org/10.1007/978-3-540-29678-2_3124.
Glickstein, M., et al. “Cerebellum: History.” Neuroscience, vol. 162, no. 3, Sept. 2009, pp. 549–559, https://doi.org/10.1016/j.neuroscience.2009.02.054.
Purves, Dale, et al. “Modulation of Movement by the Cerebellum.” Neuroscience. 2nd Edition, 2001, www.ncbi.nlm.nih.gov/books/NBK11024/.
“What Is Machine Learning? | How It Works, Techniques & Applications.” Www.mathworks.com, www.mathworks.com/discovery/machine-learning.html#:~:text=Machine%20learning%20algorithms%20use%20computational.
Singla, Sonia. “Learning the Basics of Deep Learning, ChatGPT, and Bard AI.” Analytics Vidhya, 25 Feb. 2023, www.analyticsvidhya.com/blog/2023/02/learning-the-basics-of-deep-learning-chatgpt-and-bard-ai/#:~:text=How%20is%20ChatGPT%20programmed%3F. Accessed 29 Jan. 2024.